โดย ดร. พนิดา บุญฤทธิ์ธงไชย
หลักสูตรเทคโนโลยีหลังการเก็บเกี่ยว มหาวิทยาลัยเทคโนโลยีพระจอมเกล้าธนุรี
พืชหรือสัตว์ล้วนต่างต้องเข้าสู่การเสื่อมสลายไม่ช้าก็เร็วตามแต่ลักษณะทางพันธุกรรมที่กำหนดอายุขัยของสิ่งมีชีวิตนั้น ๆ การเกิดเซลล์ตายอาจแบ่งเป็นหลายแบบ เช่น necrosis หมายถึง การที่เซลล์ตายในลักษณะเสื่อมสลายหรือแห้งตายแบบไม่มีการควบคุม เป็นการตายแบบที่เซลล์โดนทำลายโดยสิ่งที่เป็นอันตรายอย่างรุนแรงจากภายนอก เช่น สารเคมี ทางกล การอักเสบ บาดเจ็บหรืออักเสบจากบาดแผล ซึ่งเป็นผลกระทบโดยตรงต่อสาเหตุการตายนั้นๆ โดยอาการที่เกิดขึ้นในระดับเซลล์ได้แก่ เซลล์จะบวม ผนังเซลล์จะถูกทำลาย พบการแตกสลายของส่วนประกอบต่างๆภายในเซลล์ และมีการย่อยสลายของ DNA จนหมด และการตายอีกแบบหนึ่งที่จะกล่าวถึงในที่นี้ได้แก่ programmed cell death (PCD) หรือการตายโดยกำหนด การตายแบบโปรแกรม หมายถึงการที่เซลล์ตายอย่างมีรูปแบบ เซลล์นั้นๆ จะทำลายตัวเอง เมื่อโดนสิ่งกระตุ้นต่างๆ เช่น สารเคมี อุณหภูมิ เชื้อจุลินทรีย์ เป็นต้น ส่งผลให้ตัวมันเองมีกระบวนการทำลายตัวเอง เช่น การตอบสนองของพืชต่อการรุกรานของเชื้อจุลินทรีย์ซึ่งจะพบว่า บริเวณที่พืชถูกเชื้อโรคเข้าทำลาย ในส่วนรอบๆของเชื้อจะพบการตายของพืชโอบล้อมเชื้อไว้ ทำให้การแพร่กระจายของเชื้อไม่สามารถเกิดขึ้นได้ ปฏิกิริยานี้เป็นที่รู้จักกันว่า hypersensitive response (HR) การตอบสนองนี้จะจำกัดการแพร่กระจายของเชื้อโดยการที่ตัวพืชเองเกิดการตายของเซลล์พืชแบบ PCD ส่วนเซลล์ตายในสัตว์ เราจะเรียก programmed cell death ว่า apoptosis (Pennell and Lamb, 1997) ในช่วงหลายปีที่ผ่านมาการศึกษา PCD ของพืชได้รับความสนใจเป็นอย่างมากในต่างประเทศ และมีงานวิจัยจำนวนมากได้พยายามพิสูจน์ว่า การตายของเซลล์พืชที่เกิดจากสิ่งแวดล้อมภายนอก และเชื้อจุลินทรีย์ กระตุ้นให้เซลล์เกิดกระบวนการทำลายตัวเอง
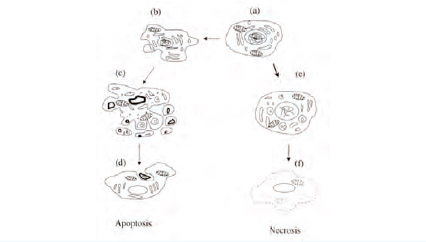
รูปที่ 1 ความแตกต่างทางสัณฐานวิทยาของเซลล์ที่มีการตายแบบ apoptosis และ necrosis (Studzinski, 1999)
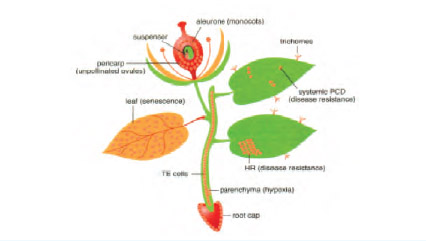
รูปที่ 2 บริเวณการเกิด programmed cell death ในพืช (Pennell และ Lamb, 1997)
กระบวนการเกิด PCD ได้แก่ DNA laddering โดย DNA จะถูกย่อยให้มีขนาด 180 bp ซึ่งเป็นลักษณะเฉพาะของเซลล์พืชและสัตว์ ที่เกิด PCD นอกจากนี้ยังการพบการทำงานของกลุ่มเอนไซม์ serine/cysteine proteinase สูงขึ้น โดยจะเกิดขึ้นในสิ่งมีชีวิตที่มีหลายเซลล์ เช่น ในสัตว์และพืชซึ่งจะกำจัดเซลล์แปลกปลอม โดยจะเกิดในช่วงการพัฒนาของเซลล์ในช่วงการพัฒนาการเจริญเติบโต (development) ซึ่งจะมีการตอบสนองต่อการติดเชื้อจากแบคทีเรีย รา และไวรัส จากภายนอก โดยสร้างสารบางชนิดส่งเข้าไปใน เชื้อโรคต่าง ๆ ส่งผลให้เซลล์เชื้อโรคเหล่านั้นทำลายตัวเอง ตัวอย่าง caspase เป็นเอนไซม์ที่ย่อยสลายโปรตีน อยู่ในกลุ่ม protease จะทำหน้าที่ในการเริ่มต้นและทำให้เกิดกระบวนการ PCD ในสัตว์
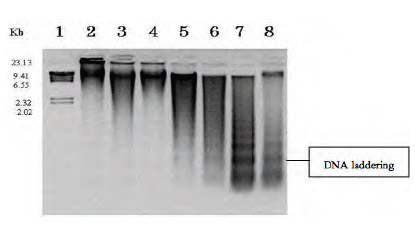
รูปที่ 3 การตรวจสอบ DNA laddering ใน DNA ที่สกัดได้จากใบ Lycoris ที่ผ่านการให้ความร้อนกระตุ้นให้เกิดการตายแบบ PCD (Boonyaritthongchai et al., 2008)
นักวิทยาศาสตร์จากมหาวิทยาลัย California, Riverside ได้ศึกษาเอนไซม์สำคัญชนิดหนึ่งที่ช่วยกระตุ้นการควบคุมกระบวนการตายของเซลล์ เพื่อป้องกันการติดเชื้อจากแบคทีเรีย รา และไวรัสของพืช และสัตว์ โดยการติดเชื้อเหล่านี้จะมีผลต่อผลผลิตของพืชอย่างมากและเป็นอันตรายต่อสัตว์บางชนิดได้ ในปี 1999 ได้มีการค้นพบว่า พืชสามารถสร้างสารพิษ ประเภท osmotin โดยโปรตีนดังกล่าวจะทำให้เซลล์ของเชื้อราเกิดการทำลายตัวเอง เพื่อช่วยหยุดการเข้าทำลายของ เชื้อรา และเป็นระบบป้องกันของพืชอีกด้วย กิจกรรมของเอนไซม์ในพืชถูกค้นว่ามีความคล้ายคลึงกับเอนไซม์ caspase ในสัตว์ซึ่งพบในช่วงขั้นตอนเริ่มแรกของกระบวนการ PCD ในขณะที่ยังไม่มีรายงานการตรวจพบยีนที่ควบคุมกิจกรรมของเอนไซม์เหล่านั้น Dr. Natasha Raikhel รายงานว่าโปรตีนที่สำคัญของพืช ได้แก่ vacuolar processing enzymes (VPEg) ในพืชทดสอบ Arabidopsis thaliana โดย VPEg ช่วยป้องกันการติดเชื้อจากแบคทีเรีย รา และไวรัส ที่ก่อให้เกิดโรคในพืช โดยจะไปกระตุ้นระบบ PCD ของเชื้อโรค โดยการควบคุมกระบวนการตายของเซลล์ที่เป็นสาเหตุสำคัญในกระบวนการที่เกี่ยวข้องกับการเปลี่ยนแปลงทางสรีระวิทยามากมาย โดยมี VPEg เป็นตัวควบคุม PCD
PCD เป็นกระบวนการที่เกิดขึ้นในระดับชีวโมเลกุลที่มีความเกี่ยวข้องกับการเปลี่ยนแปลงลักษณะทางสัณฐานวิทยาและกระบวนการทางชีวเคมี โดยการเปลี่ยนแปลงในระดับชีวโมเลกุลนี้จะเกิดขึ้นอย่างรวดเร็ว เช่น การเพิ่มขึ้นของ ออกซิเจนในรูปแอคทีฟ (reactive oxygen species) การทำงานของเอนไซม์ caspase ซึ่งเป็นเอนไซม์ที่สำคัญของการเกิด PCD และการเกิด DNA laddering ที่มีขนาด 180 bp เป็นต้น ซึ่งการเปลี่ยนแปลงทางชีวโมเลกุลนี้จะส่งผลต่อ การเปลี่ยนแปลงทางกายภาพในภายหลัง เช่นการเปลี่ยนแปลงของสีผิวและสีเปลือก ลักษณะการอ่อนนุ่มของผนังเซลล์ และเป็นที่ยอมรับกันว่า PCD เป็นส่วนที่สำคัญที่เกิดขึ้นภายในช่วงชีวิตของสิ่งมีชีวิตทั้งหลาย ทั้งพืชและสัตว์ สำหรับพืช PCD จัดเป็นกระบวนการที่สำคัญสำหรับการพัฒนาและการมีชีวิตของเซลล์พืชรวมทั้งการป้องกันตัวจากสิ่งกระตุ้นจากภายนอก นอกจากนี้ PCD ยังเกี่ยวข้องการเสื่อมสภาพของพืช (senescence) อีกด้วย การเสื่อมสภาพหรือการชราภาพของผลิตผลหลังการเก็บเกี่ยวจะพบการสลายตัวของคลอโรฟิลล์เป็นการเปลี่ยนแปลงที่สามารถสังเกตเห็นได้ชัดเจนที่สุด และมักจะเกิดขึ้นพร้อมๆ กับการสูญเสียไขมันและโปรตีนที่เป็นองค์ประกอบของเยื่อหุ้มต่าง ๆ และท้ายที่สุดส่งผลทำให้เซลล์พืชตาย การเสื่อมสภาพของพืชจัดเป็นกระบวนการที่ตอบสนองต่อสัญญาณการกระตุ้นทั้งจากภายนอกและภายใน ซึ่งต้องอาศัยการแสดงออกของยีน (gene expression) การสังเคราะห์โปรตีน (protein synthesis) และการเสื่อมสภาพนี้มีความเกี่ยวข้องกับกระบวนการชีวเคมีต่าง ๆ ที่ควบคุมโดย signaling pathway (Buchanan- Wollaston, 1997; Chandlee, 2001) และมีความเกี่ยวข้องกับ PCD (Gan and Amasino, 1997) Coupe et al. (2003) พบว่า การเสื่อมสภาพของบลอคโคลี่ภายหลังการเก็บเกี่ยวมีความเกี่ยวข้องกับการทำงานของเอนไซม์พวก cysteine protease ซึ่งเป็นเอนไซม์ที่เกี่ยวข้องกับการตายแบบ PCD นอกจากนั้น Eason et al. (2002) พบ DNA laddering ในหน่อไม้ฝรั่งที่เสื่อมสภาพภายหลังการเก็บเกี่ยว ซึ่งเป็นลักษณะอาการของ PCD นอกจากนี้ การศึกษา PCD ในพืชพบว่าที่อุณหภูมิ 55ºC เป็นเวลา 10 นาที สามารถกระตุ้นให้พืชแสดงอาการ PCD ได้ Vacca et al., 2004 แต่อย่างไรก็ตามความสัมพันธ์ระหว่างการเกิด PCD ต่อการเสื่อมสภาพของผลิตผลทางการเกษตรภายหลังการเก็บเกี่ยวยังไม่เป็นที่ทราบแน่ชัด เพราะฉะนั้นการศึกษาเรื่องการสูญเสียของผลิตผลภายหลังการเก็บเกี่ยวและการเกิด PCD จึงมีความสำคัญ เพื่อที่จะหาวิธีป้องกันการสูญเสียที่เพิ่มมากขึ้นภายหลังการเก็บเกี่ยวของผลิตผลทางการเกษตร และการศึกษาเรื่อง PCD ในพืชนับเป็นเรื่องที่ใหม่และมีการตื่นตัวในการศึกษาวิจัย อันจะเป็นประโยชน์ต่อไปในอนาคต
เอกสารอ้างอิง
- Pennell, R. I. and C. Lamb. 1997. Programmed cell death in plants. The Plant Cell. 9: 1157-1168.
- Studzinski, G. P. 1999. Overview of apoptosis. In: Apoptosis A Practical Approach. Studzinski GP (eds), Oxford University Press, New York. 1-17.
- Boonyaritthongchai, P., Manuwong, S., Kanlayanarat, S., and T. Matsuo. 2008. Accelerated of senescence by high temperature treatment in Lycoris (L. traubii x L. sanguinea) leaf sections. Journal of the Japanese Society of Horticultural Science. 77(4): 431-439.
- Buchanan-Wollaston, V. 1997. The molecular biology of leaf senescence. J. Exp. Bot. 48: 181-99.
- Chandlee, J.M. 2001. Current molecular understanding of the genetically programmed process of leaf senescence. Physiologia Plantarum. 113: 1-8.
- Gan, S. S. and R. M. Amasino. 1997. Making sense of senescence molecular genetic regulation and manipulation of leaf senescence. Plant Physiology. 113: 313-319.
- Coupe, S. A., B. K. Sinclair, M. Watson, J. A. Heyes and J. R. Eason. 2003. Identification of dehydration-responsive cysteine proteases during postharvest senescence of broccoli florets. Journal of Experimental Botany. 54:1045-1056.
- Eason, J. R., T. T. Pinkney and J. W. Johnston. 2002. DNA fragmentation and nuclear degradation during harvest-induced senescence of asparagus spears. Postharvest Biology and Technology. 26: 231-235.
- Vacca, R. A., M. C. de Pinto, D. Valenti, S. Passerella, E. Marra, and L. De Gara. 2004. Reactive oxygen species production, impairment of glucose oxidation and cytosolic ascorbate peroxidase are early events in heat-shock induced programmed cell death in tobacco BY-2 cells. Plant Physiology. 134: 1100–1112.
บทความนี้ ตีพิมพ์ใน Postharvest Newsletter ปีที่ 10 ฉบับที่ 3 กรกฎาคม – กันยายน 2554


